Replikacija, dvigubėjimas
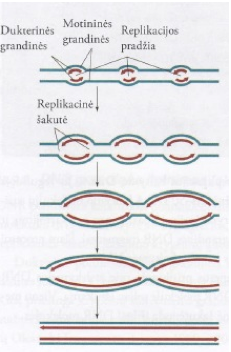
Tai – DNR sintezė. DNR dvigubėjimas (replikacija) vyksta pusiau konservatyviuoju mechanizmu. Jo metu iš vienos DNR molekulės susidaro dvi vienodos DNR molekulės. Susidariusios naujos DNR molekulės viena polinukleotidinė grandinė yra sena (motininė), o kita (dukterinė) sintetinama nauja. 1953 m. J. Votsonas ir F. Krikas pasiūlė DNR dvigubosios spiralės modelį, kuris paaiškino DNR dvigubėjimo principą: vienoje polinukleotidinėje grandinėje bazės yra komplementariosios kitoje grandinėje esančioms bazėms. Vykstant biosintezei, komplementariosios DNR grandinės atskiriamos ir fermentai DNR polimerazės katalizuoja dukterinių DNR molekulių sintezę.

Dvigubėjimo (replikacijos) etapai: 1) pradėtis (iniciacija) prasideda, prisijungus dvigubėjant (replikacijoje) dalyvaujantiems baltymams prie DNR molekulės; 2) ilginimo (elongacijos) metu DNR polimerazė katalizuoja naujų nukleotidų įsiterpimą į ilgėjančią polinukleotidinę grandinę; 3) baigties (terminacijos) metu DNR biosintezė sustoja, nuo DNR atskyla biosintezėje dalyvaujantys komponentai ir dvi naujos DNR molekulės atsiskiria.
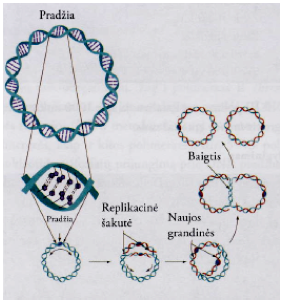
Dvigubėjimo (replikacijos) etapai:
Pradėtis – DNR dvigubėjimas (replikacija) prasideda savitajame DNR pradžios taške. E. coli jis yra vadinamas oriC. Pradėties pradžios taškas sudarytas iš 245 bazių porų.
Dvigubėjant E. coli DNR dalyvaujantys baltymai: baltymas DnaA. Jis reguliuoja DNR dvigubėjimą (replikaciją), kontroliuodamas pradėties dažnį. 20–50 DnaA baltymo molekulių susijungia su daug A-T porų turinčia seka, kuri sudaryta iš trijų 13 bazių porų pasikartojimų, ir ją išlydo. Dvi grandinės atsiskiria ir susidaro viengrandės DNR fragmentai. Šiam procesui panaudojama ATP energija ir dalyvauja į histonus panašus baltymas HU. Fermentas helikazė (DnaB baltymas) prisijungia prie atsiskyrusios DNR grandinės. Naudojant ATP hidrolizės energiją, DNR molekulė toliau išsukama į abi puses nuo ori taško. SSB baltymai jungiasi prie išsuktos viengrandės DNR molekulės. Jie stabilizuoja DNR grandinę, apsaugo ją nuo renatūracijos ir nuo nukleazių, hidrolizuojančių viengrandę DNR, poveikio, neleidžia susidaryti dvigubajai spiralei. DNR topoizomerazės hidrolizuoja fosfodiesterinį ryšį ir nutraukia DNR grandinę. I tipo DNR topoizomerazė skelia vieną dvigubosios DNR molekulės grandinę ir vėliau jas sujungia. Fermentas turi nukleazinį (hidrolizuoja fosfodiesterinius ryšius) ir ligazinį (sujungia grandines) aktyvumą. II tipo DNR topoizomerazės tvirtai susijungia su DNR dvigubąja spirale, sukelia trumpalaikį trūkį abiejose grandinėse, pašalina teigiamąją ir neigiamąją superspiralizacijas.
Eukariotų chromosomos dvigubėjimas (replikacija) prasideda vienu metu daugelyje taškų, kurie nutolę vienas nuo kito per 30–300 kbp ir vyksta į abi puses. Viena eukariotų chromosoma turi daug dvigubėjimo (replikacijos) pradžios taškų. Dvigubėjimo pradžios sritis yra vadinama ARS (savarankiškai dvigubėjanti seka), arba replikonu. Dvigubėjimo (replikacijos) pradžiai taip pat reikia baltymo ors, kuris susijungia su atitinkamomis replikatoriaus sritimis. Eukariotų chromosomos dvigubėja per 1 valandą.
Ilginimas – DNR polinukleotidinės grandinės ilginimas vyksta 5’→3′ kryptimi. DNR molekulės dvi grandinės yra antilygiagrečios, todėl vienos naujos grandinės sintezė sutampa su replikacinės šakutės judėjimo kryptimi, kitos grandinės sintezė turi vykti priešinga kryptimi. Dukterinė polinukleotidinė grandinė, kurios sintezės kryptis sutampa su replikacinės šakutės judėjimo kryptimi, sintetinama nenutrūkstamai ir yra vadinama pirmaujančiąja. Kita, priešinga kryptimi sintetinama grandinė, vadinama vėluojančiąja. Jos sintezė vyksta mažais fragmentais, vadinamais Okazaki fragmentais. Prokariotų Okazaki fragrnento ilgis yra 1000–2000 bp, o eukariotų – 100–400 bp. Polinukleotidinės grandinės ilginimą katalizuoja DNR III polimerazė. Prie pradmens ribonukleotido 3′-galo laisvos hidroksigrupės prijungiami deoksiribonukleotidai ir pirmaujančioji grandinė nuolatos ilgėja. Vėluojančiosios grandinės sintezė prasideda taip pat nuo RNR pradmens, tačiau vyksta trumpais Okazaki fragmentais ir priešinga kryptimi. Helikazė ir praimazė sudaro funkcinį dvigubėjimo (replikacijos) komplekso vienetą, vadinamą praimosoma.
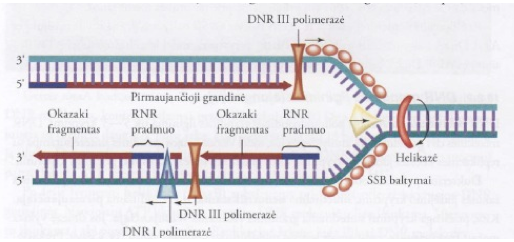
Susidarę Okazaki fragmentai yra sujungiami (pašalinus RNR pradmenį ir vietoj jo susintetinus DNR) ir susidaro nenutrūkstama polinukleotidinė grandinė. Šias reakcijas katalizuoja fermentai DNR I polimerazė ir DNR ligazė. Eukariotų pirmaujančioji DNR grandinė sintetinama nenutrūkstamai, o vėluojančiosios grandinės sintezė vyksta fragmentais. Vėluojančiosios grandinės sintezė vyksta atskirais etapais, kaip ir E. coli (pradmens sintezė, Okazaki fragmentų sintezė, pradmens hidrolizė, tarpų tarp polinukleotidinių fragmentų užpildymas, pavienių fragmentų sujungimas). Daugelis eukariotų ląstelių turi mažiausiai dešimt skirtingų DNR polimerazių. Eukariotų DNR susijungia su histonais ir sudaro nukleosomas, tai gali lemti lėtesnį replikacinės šakutės judėjimą. DNR dvigubėjimas (replikacija) yra susijęs su tuo pačiu metu vykstančia histonų biosinteze. Nauji histonai jungiasi su dukterinėmis DNR polinukleotidinėmis grandinėmis.
Baigtis – E. coli dvigubėjimo (replikacijos) baigtis vyksta baigties taške Ter (lot. terminus –baigtinis), kuris išsidėsto žiedinės chromosomos priešingoje ori taškui pusėje. Šioje srityje DNR molekulėje yra daug 20 bp savitųjų bazių kopijų, kurios susijungia su baltymu Tus (angl. termination utilization substance). Baltymo molekulėje yra ryškios β strukūros. Jos susijungia su DNR molekulės didžiuoju grioviu. Baltymas slopina replisomos helikazinį aktyvumą ir replikacinės šakutės judėjimą. Baigties srityje taip pat yra nukleotidų sekos, kurios palengvina dukterinės chromosomos atsiskyrimą, pasibaigus DNR dvigubėjimui (replikacijai). Į eukariotų chromosomos dvigubėjimo (replikacijos) baigtį įtraukiamos specialios struktūros, vadinamos telomeromis, kurios prijungiamos DNR molekulės gale ir stabdo replikacinės šakutės judėjimą.
Replikacinė šakutė – replikacijos metu dvi DNR grandinės išsiskiria. Susidaro V raidės formos struktūra, kuri vadinama replikacine šakute. Replisoma prisijungia prie kiekvienos iš dviejų replikacinių šakučių.

Šaltinis | Dažniausiai vartojamų biomedicinos terminų ir sąvokų aiškinamasis žinynas | Lietuvos sveikatos mokslų universitetas | Akademikas Profesorius Antanas Praškevičius, Profesorė Laima Ivanovienė